
摘要
本研究试图检验利用浮游植物作为生物指标评价埃及东亚历山大海岸水质的适用性。选择了8个站点来覆盖2022年夏季海水的不同特征。以不同比例检出了硅藻门、蓝藻门、藻门、绿藻门、硅鞭毛门和裸藻门6个藻类类群。塞得港(PS)海滩浮游植物丰度最高,Ras El Bar (RB)海滩最低。根据浮游植物丰度评价,除PS为富营养化外,其余试验站均为贫营养化状态。根据Chl a浓度,所选站点从西到东表现为好-坏。Shannon多样性指数和Pielou均匀度指数将研究站点划分为中度污染,除了PS和RB站点。Navicula、Synedra、Euglena等有机污染指标物种的存在是水质下降的预警信号,特别是在B站和M站。采集到的浮游植物的生化成分具有空间波动特征。与脂类相比,蛋白质和碳水化合物是浮游植物群落中占优势的大分子。这些大分子用于评估营养状态,而在一些站点观察到的蛋白质和碳水化合物之间的比率> 1代表健康海岸。因此,浮游植物应被视为地中海水框架指令监测计划中的生物指标。为了对地中海生态系统进行长期监测,建议对浮游植物的大分子进行测定。
介绍
地中海是一个众所周知的贫营养水体,它缺乏营养而富含溶解氧。营养不良的状况从西向东蔓延。在过去几年中,由于水性质的变化,这种模式可能发生了变化,这可能是人类活动的结果(Christaki et al., 2011)。城市化和旅游业造成沿岸污水污染,提高了东部和西部盆地的海水温度和盐度(Bethoux & Gentili, 1999)。值得注意的是,联合国环境规划署(UNEP)报告称,每年有6.5亿吨污水排入地中海(Karadirek et al., 2019)。除了地中海沿岸埃及持续的建筑活动、快速的工业发展和人口的不断激增外,该地区的海水污染日益严重,成为东地中海污染最严重的地区之一(Heger et al., 2022)。埃及地中海沿岸最重要的地区之一是亚历山大沿海地区,那里有许多海滩。它从西部的El-Dekhaila延伸到东部的Abu Qir (Dango et al., 2015)。另一方面,Ras El-Bar位于地中海Damietta分支的河口,尼罗河的淡水与地中海的盐水在这里混合。Damietta河口接收来自Ras El-Bar污水处理厂等污水处理设施、农业径流、Moboco化肥厂等工业作业以及Ezbt Elborg的繁忙船只交通的排放物(Abdel Galil等,2020)。巴尔提姆海滩是尼罗河三角洲中心部分最重要的公共海滩之一,位于埃及北岸,位于布鲁勒斯泻湖入口以东11.5公里处。此外,塞得港海岸还受到两个重要水体的影响,即苏伊士运河和曼萨拉湖,导致地表水主要来自地中海。大量文献表明,相当数量的赤丝瓜物种已经通过苏伊士运河迁移,入侵地中海东部(Madkour, 2000)。湖水流出引起的淡水流入程度根据该地区的盛行风模式呈现季节性变化,主要是由北到西北,风速在6至14.5节之间(Amer, 1999年)。
最近,人们似乎对地中海的多种浮游植物产生了兴趣,由于污染、船舶交通、气候变化、引入物种和本地物种分布的变化,地中海的浮游植物正在迅速变化(El-Dahhar等人,2021;Ismail et al., 2022)。
浮游植物群落在海洋生物多样性和生产力以及监测环境变化方面发挥着重要作用。然而,它们的丰度和结构会随着环境参数的变化而变化,因此它们可以用作水质和富营养化程度的生物指标(Kim et al., 2020)。由于生物和非生物环境条件对海洋环境中浮游植物的演替和丰度都有重大影响,这些变化在海洋生态系统中多重营养循环的耦合中起着巨大作用。然而,浮游植物组合的波动是由许多因素解释的,如养分供应、光照变化和混合条件(Litchman et al., 2015)。因此,浮游植物的丰度、物种组成、丰富度、均匀度以及地理和时间分散反映了给定水体的生物完整性或环境福祉(Effiong et al., 2018)。此外,一些指示藻类物种是水污染的关键指标(Atıcı & Akiska, 2005)。
此外,浮游植物是海洋生态系统中碳的主要主要生产者,随后在浮游植物生物量中产生许多必需的生物分子,如总碳水化合物、蛋白质和脂质,这些生物分子具有潜在的高度生物活性(Heraud等人,2008;Ismail et al., 2022;Kim等人,2022),并且可以有效预测其营养价值(Ahn等人,2019;Ismail et al., 2022)。主要生化成分的变化多受生长阶段、物种组成和环境参数控制(Ahn et al., 2019;Ismail et al., 2022)。较高营养水平的食物质量受到浮游植物生化组成变化的显著影响(Kim et al., 2022)。
一些当地研究集中在东亚历山大海岸一个或多个地区的浮游植物群落结构上(El-Sayed等人,2019;El-Serehy et al., 2014;加里布和多格姆,2000;侯赛因,2008;伊斯梅尔和易卜拉欣,2017;Labib et al., 2023)。
本文研究了东埃及地中海沿岸从西(东港)到东(塞得港)的浮游植物群落组成及其大分子组成(碳水化合物、蛋白质和脂质)与理化参数的空间变化。本研究还通过使用多种统计方法和指数,为地中海沿岸环境的管理研究提供了有用的见解。
材料与方法
采样地点
2022年初夏,研究人员从亚历山大东部海岸八个地点收集了水和浮游植物样本,这些地点覆盖了受自然和/或人为干扰影响的不同生态实体。这些网站是东部港口(呃),Sidi Gaber (SG) Gleem (G),阿布Qir (AQ) Boughaz El Maadiya (M), Baltim (B), Ras El酒吧(RB)和端口(PS)表示(图1和表1)。电台嗯和PS是商业港口受到不同的人为活动,比如污水和航运,以及电台AQ和B,所影响的大量未经处理的污水,农业,工业,和污水,除了现有的贸易船只在RB车站。G站和SG站代表未受污染的地区,因为它们受自西向东的水流影响。El Maaddiya海峡大约100米长,20米宽,3米深,是Abu Qir湾和Idku湖的水交换的地方。由于地中海海水与艾德库湖的微咸水不断混合,该地区受到不规则变化的影响,这造成了不稳定的生态特征(Gharib和doghham, 2000)。此外,它还接收来自Edku湖的排水,其中含有来自Beheira省和其他地区的农业、工业和城市化学物质,含量不详。
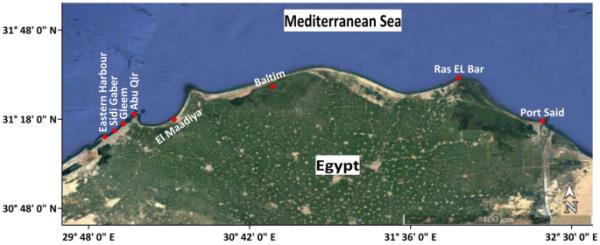
选定车站的地图
海水的理化参数
水温、盐度、溶解氧和pH的表面测量使用水检物理参数装置(HANA,型号HI 9828)进行。溶解的无机营养物(NO2、NO3、NH4、PO4和SiO4)的分析参照Parsons et al.(1984)的方法,叶绿素a (Chl a)的估算参照Parsons(2013)的方法。
浮游植物收集与鉴定
地表水样品使用细网(20µm网)和垂直拖网(0.5 m/s)获得,然后使用流量计(HYDRO-BIOS, Kiel,德国)测定过滤后的水。收集的样品保存在中和的福尔马林(4%)中,然后在黑暗中储存几滴Lugol酸溶液,然后运往埃及亚历山大水生生物群分类和生物多样性实验室进行分类检查和计数。浮游植物分析使用倒置显微镜(Optika 100)进行,沉淀后放大倍数为400倍(Utermöhl, 1958)。浮游植物的分类鉴定是根据Krammer and Lange-Berlatot(1991)和center -Lund and Lund(1996)进行的,然后通过藻类基地网站进行确认(Guiry & Guiry, 2022)。
不同指标对水质的评价
有不同的测定方法用于估计水质,包括浮游植物丰度(Kitsiou & Karydis, 2001),叶绿素a含量指数(Karydis, 1999),并由Simboura等人(2005)修改。此外,富营养化指数(E.I.)是评估营养状态的多指标(营养物和Chl a的组合)工具(Primpas et al., 2010)。富营养化指数的计算公式如下:
在E.I. < 0.04(高生态水质),E.I. = 0.04 - -0.38(好),E.I. = 0.38 - -0.85(中度),E.I. = 0.85 - -1.51(贫穷),和E.I. > 1.51(坏的管理者)。
物种多样性(H′)(Shannon, 1997)、物种丰富度(d) (Margalef, 1978)和物种均匀度(J) (Pielou, 1975)。
其中N是所有物种的总个体数,ni是一个物种的个体数。
其中S为物种总数,N为样本中个体总数。
其中H为香农指数,S为物种数。
Palmer污染指数是根据发现的藻类类群来检测环境污染水平的(Palmer, 1969)。
相似性指数
利用Sorenson方程(Sorenson, 1948)计算研究区浮游植物物种之间的相似度作为统计参数,该相似度取决于不同物种的存在与否:
在哪里
- 空间站:
-
相似性系数
- C:
-
在两个地点共有的种数
- 答:
-
第一站点的物种数量
- B:
-
第二点的物种数量
浮游植物群落的生化组成
使用塑料瓶从选定的地点收集已知体积的海水样本,随后使用浮游动物网(100微米网目尺寸)对其进行筛选和过滤,以分离大型浮游动物。样品被移至冰槽实验室,并在Whatman GF/F(0.7-µm孔径)纤维圈上重新过滤,以测量分离后浮游植物的生化特性。总蛋白含量(PRO)采用Biuret法测定(David & Hazel, 1993)。总碳水化合物含量(CHO)采用Dubois et al.(1956)建立的方法。总脂质含量(LIP)采用Bligh和Dyer(1959)的方法检测。
统计分析
基于样本间的Jaccard和Bray-Curtis距离矩阵,采用Primer 6.1.9软件(Primer- e Ltd.)进行聚类分析,生成树形图(组平均法)。采用SPSS软件30,2020,采用单因素方差分析(One-way ANOVA)检验确定各站点之间与水和浮游植物生物量的各种理化性质相关的差异。利用Microsoft Excel 2018软件估算相关系数(r),评价研究区浮游植物的理化参数与浮游植物丰度、类群及其生化组成(n = 21)之间的关系。
目录
摘要介绍
材料与方法
结果与讨论
结论
数据可用性
参考文献
致谢
作者信息
道德声明
补充信息
搜索
导航
#####
结果与讨论
物理化学参数
表2列出了2022年夏季东亚历山大海岸水质参数的空间变化。研究站点之间的物理和营养值存在显著差异(单因素方差分析,p < 0.05)。温度的特征是站与站之间略有不同;B站最低(17℃),EH站最高(21℃)。8个研究点的pH值为微碱性,B站的pH值为7.94,RB和EH站的pH值为8.16。盐度是与浮游生物多样性相关的主要物理参数,反映了水生环境中的污染程度(Zyadah et al., 2004)。它在AQ和PS站表现出高振荡,最大值为41.20 PSU,超过了地中海水域(38.50 PSU) (copin - mont
一般来说,海水的特点是高水平的营养物质,表明排放的水到达的影响。总氮解释了NO2、NO3和NH4的浓度。硝酸盐是无机氮化合物中含量最高的组分,其浓度从AQ最低(0.19µM)到G站最高(18.78µM)。AQ站和M站亚硝酸盐浓度分别在0.15 ~ 5.55µM之间。此外,不同站点之间的浓度差异很大,M站和B站的浓度分别为0.04 ~ 8.12µM。大多数监测站的NO3(4µM)和NH4(2µM)水平是富营养化的标准(Oczkowski & Nioxn, 2008)。AQ站和PS站的磷酸盐浓度分别在0.24 ~ 5.62µM之间波动。根据Stirn(1988)的报道,在被污染的水中,通常存在大量的可溶性磷,因此与其他选定的站点相比,PS, m和RB被认为是高污染区域。EH、G、AQ和B 4个站点的N:P比值均高于Redfield站点(16:1),显示出较高的氮收支,说明磷在这些站点发挥了限制作用。SiO4浓度在不同位点之间表现出较高的振荡,PS位点的峰值为10.72µM, AQ位点的峰值较低(1.60µM)。这个宽范围与硅藻门(硅藻)的丰度有关(r = 0.61)(表2S)。Chl a(平均1.22µg L−1)的空间变异性较大,p < 0.05。B和RB有两个明显的峰(> 1µg L−1),并伴有浮游植物快速增殖(r = 0.74)。然而,随着海水中磷酸盐和硝酸盐含量的增加,Chl a和Chl a都会增加(Nassar & Hamed, 2003)。这与目前的研究结果一致,Chl-a与磷酸盐(r = 0.719)和硝酸盐(r = 0.616)呈显著正相关(p≤0.05)。一般来说,水的物理化学性质的区域差异可归因于排放的废水,如排水和捕鱼。特别是,营养物(硝酸盐和磷酸盐)的高浓度可归因于某些站点的人为活动的影响,如捕鱼、游泳、农业废物、工业废物和污水废物。本研究中理化参数的总体平均值与前人在亚历山大海岸观测到的结果或多或少相似(Labib et al., 2023;Alprol et al., 2021)。
浮游植物群落的结构和组成
研究站共鉴定出126种藻类。一般来说,记录的物种可以被认为是温带水的形式,泛热的,以前在地中海被报道过。硅藻丰富度指数最高(35属,87个类群),丰度最高(占总丰度的91.06%),鞭毛藻(Dinophyta) 14属,30个类群)丰度次之(1.81%)。蓝藻科有4种。绿藻门和硅鞭毛门均为2种,裸藻门为1种(表1S)。与以往的研究比较,发现与Alprol等人(2021)和Dango等人(2015)或多或少相似。总的来说,除了B站之外,硅藻物种在所有站点中都是优势种群,因为它们对包括极端条件在内的不同水域的生存具有高度的适应性(Odum, 1998),因此它们可以用作未污染水域的生物指示物(Al-Tamimi & Al-Jumaily, 2021)。硅藻的优势度受Si (r = 0.613)和PO4 (r = 0.555)的控制,因为PO4和SiO4是控制浮游植物生长的主要因素(Rollwagen-Bollens & Connelly, 2022)。B站蓝藻密度最大,主要是淡水藻类“铜绿微囊藻(k
浮游植物的分布和丰度
研究区浮游植物丰度和生物多样性存在显著的空间差异(单因素方差分析,p < 0.05)。这种变化取决于物理和化学参数的相互作用,而这些参数又受到气候和人为活动的影响(Kim et al., 2020)。理化参数与浮游植物丰度呈正相关,反映了其在浮游植物生物量空间变化中的重要作用。一般而言,浮游植物群落丰度与硅藻(r = 0.996)和鞭毛藻(r = 0.989)显著相关,它们被认为是海洋环境中浮游植物的主要组成部分(Limates et al., 2016)。此外,铵在浮游植物丰度中起着至关重要的作用(r = 0.76),因为NH4是浮游植物首选的氮源(Straskraba & Tundisi, 1999)。除了水体的物理化学参数外,还有许多其他因素可以影响浮游植物的空间分布,如浊度最大位置和大小、潮汐振幅和淡水排放量的相对重要性、水柱分层和浮游动物放牧率(Harrison et al., 1991)。
浮游植物丰度平均最高的是PS海岸(1.77 × 106细胞L−1),其次是B (177 × 105细胞L−1)、M (963 × 104细胞L−1)、AQ (683 × 104细胞L−1)、SG (646 × 104细胞L−1)和G (532 × 104细胞L−1),如图2所示。EH (420 × 104)和RB (375 × 104细胞L−1)浮游植物丰度最低。PS站和B站的浮游植物丰度最高,分别与骨骨藻(1.7 × 106细胞L−1)和铜绿微囊藻(1.5 × 105细胞L−1)的过度生长有关。
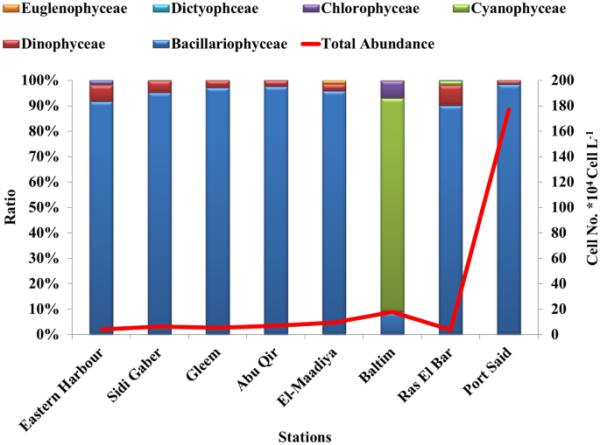
浮游植物群落丰度(细胞数)的空间变异。× 104 cell L−1)和ratio %
测定的浮游植物生物量的范围与之前在亚历山大不同海岸记录的浮游植物生物量的范围相似(236.4 × 103 cells L−1)(Zaghloul, 1994)和(680 × 103 cells L−1)(Hussein, 2008)。结果表明:EH站中肋骨藻占主导地位(35.5%),SG站占主导地位(20.7%),G站占主导地位(39.8%),AQ站占主导地位(28.3%),M站、RB站和PS站中肋骨藻占主导地位(分别为31.9%、52.7%和96%),B站中铜绿微囊藻占主导地位(82.5%)。
根据浮游植物细胞密度评价(Kitsiou & Karydis, 2001),除PS(丰度> 10 × 105)为富营养状态外,东地中海水体所选站点均处于贫营养状态(丰度< 3 × 105)。
空间消长上优势种差异较大。这包括骨骨藻(Greville) Cleve,它构成了除B站外大多数站点浮游植物丰度的主体。此前,S. costatum已被确定为埃及地中海水域Damietta分支口(Halim, 1960)、东部港口(Ismail & Ibrahium, 2017)和Abu-Qir湾(El-Sherif & Mikhail, 2003)中偶尔引起红潮的开花物种。此外,海岸带骷髅因其高的耐污染能力而成为富营养化的指示物种(Nassar & Hamed, 2003)。大多数站点的优势硅藻为细粒藻(Melosira granulata (Ehrenberg) Ralfs)、不同毛藻(不同点)、Pleurosigma rigum W. Smith和pseudonitzschia pungens (Grunow ex Cleve) Hasle。来自蓝藻门的铜绿微囊藻k
叶绿素一个浓度
叶绿素a浓度本身可以作为确定水体营养状况的指导。PS中Chl a含量最高,为3.24µg L−1。EH海滩Chl a含量最低,为0.27µg L−1。
总的来说,所有选择站点的Chl a估估值都低于Alprol等人(2021)在地中海东南部亚历山大的8个站点检测到的6.96µg L−1。根据Karydis(1999)提出的、Simboura等人(2005)修改的基于Chl a浓度的希腊海富营养化规模,也适用于《水框架指令》定义的五个生态状况级别。而富营养化规模则根据Chl a分为优质水系统(< 0.1µg L−1)、低营养(良好)(0.1 ~ 0.4µg L−1)、中营养(中等)(0.4 ~ 0.6µg L−1)、差(0.6 ~ 2.21µg L−1)和坏(> 2.21µg L−1)。从表3可以看出,西部(EH、SG、G、AQ站)水质为好/中,东部(M、B、RB、PS站)水质为差。
Chl A与浮游植物丰度呈显著正相关(r = 0.74),特别是硅藻(r = 0.69)和鞭毛藻(r = 0.65),这可能解释了这些物种在研究区域的局限存在。Chl a与NO2 (r = 0.616)、PO4 (r = 0.719)呈正相关(p < 0.05)。
富营养化指数(E.I.)
计算出的E.I.值在1.5以上,表明富营养化程度普遍存在,B站的E.I.值最大,为8.13,AQ站的E.I.值最小,为1.43,表明所有研究区域都处于营养不良状态(表3),反映了海洋系统发生了显著的生态变化(Primpas et al., 2010;Simboura et al., 2016),其原因可能与表1中提到的不同的人为活动有关。
生物多样性指数
物种多样性、均匀度和物种丰富度等指标被估计为定义浮游植物群落结构的参数(图3)。影响多样性的因素有很多,如捕食或竞争等生物因素,以及栖息地严酷度、异质性或大小等非生物因素(Estrada et al., 2004)。已经进行了一些数值尝试,以浮游植物种类而不是营养水浓度来表示贫营养化和富营养化水平。根据Shannon分类,除B站(0.983 nats)为中度污染(2-3 nats), PS站(0.266 nats)为重度污染(Shannon et al.(1997)分类),H '值低表明污染程度高,H '指数值高表明多样性丰富。与以往在PS站观察到的铜绿假单胞菌(M. aeruginosa)在B和S. costatum的优势相同,该值较低时,其类群数量较少,且为单一或少数种的优势。
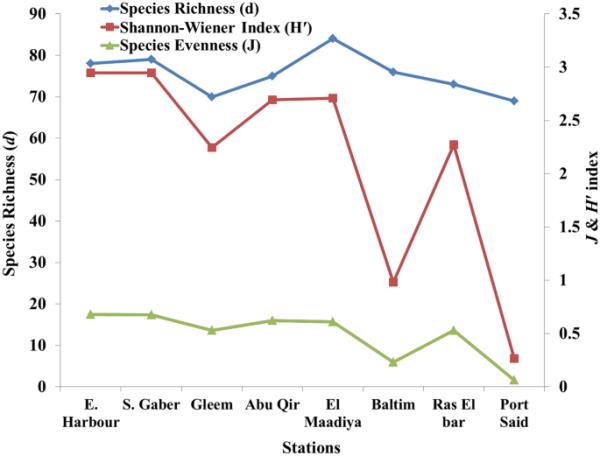
亚历山大东部沿海浮游植物群的丰富度、均匀度和香农指数
利用Pielou均匀度指数(J)对所选站点的浮游植物多样性进行评价。PS站和EH站浮游植物J指数在0.06 ~ 0.68之间,平均值为0.49,与Shannon H的结果一致。而根据国际标准分类,除PS(0.06)和B(0.23)站外,大多数选择的站点都是中度污染地区(0.5-0.6)(Zheng et al., 2007)。
如图3所示,沿东地中海和埃及检测到浮游植物丰富度的空间变化(d)。各测点的物种总数存在轻微的空间差异。El Maadiya有84种,其次是SG,共发现79种。RB站、AQ站、B站和EH站的种数比较接近(73 ~ 78种),而PS站和G站的种数较少(69 ~ 70种)。藻类群的生物多样性因水的性质而异。物种丰富度是了解群落生物多样性和动态的重要因素。在我们的研究中,除B站外,硅藻是各区域物种丰富度的重要贡献者。总体而言,检测到的丰富度低于Alprol等人(2021)(228种)和El-Dahhar等人(2021)(169种)在不同埃及地中海海岸观察到的丰富度。
帕尔默污染指数
污染指示物种是决定生物水质的关键因素。帕尔默指数是预测地表水污染的一种有效方法。帕尔默藻种类的发生和分布情况见表4。由于有许多物种可以作为生态系统质量的生物指标(Atıcı & Abel, 2016),在研究区域检测到8种耐污染藻类类群。根据浮游植物物种Palmer污染指数得分(Palmer, 1969), B站和RB站可能是有机污染,其他站Palmer指数得分< 10,没有污染证据。
空间相似性和站点分组
为了量化浮游植物种群的空间变异,我们使用了布雷-柯蒂斯相似指数。聚类分析是一种用于将相似变量分组的统计技术。在树形图方面,采样点被分为两个统计上显著的簇。第一组包括Baltim采样点和第二组的所有其他选定区域。从浮游植物群落组成的检查(图4)来看,由于所有区域都具有相同的优势群(硅藻),除了巴尔提姆外,其特征是蓝藻类的优势。
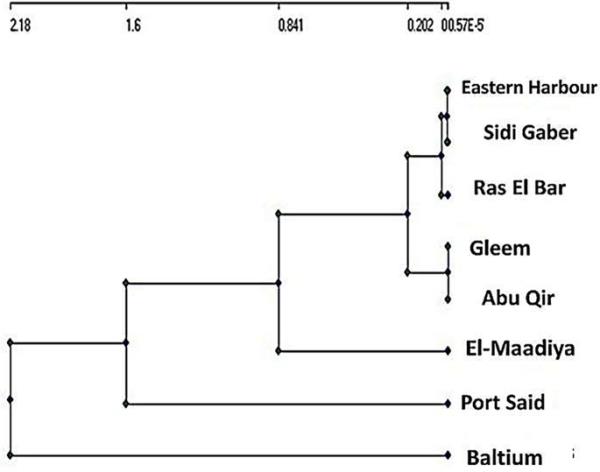
对各监测点进行聚类分析
采用Kulczynski系数相似指数测定不同站点之间藻类种类组成的相似程度(表5)。相似指数显示,B站和M站的相似度最高,为66.20%,其次是B站和PS站(60.44%),而B站和G站的相似度最低,为36.40%。这种变化与藻类种类和丰度有关。
浮游植物群落的生化组成
细胞水平生物标志物(PRT、CHO和LIP含量)的变化能够阐明污染暴露后可能伴随结构和功能改变(初级生产力和营养吸收率)的细胞生化反应(Heraud等人,2008)。生物化学模式被用作监测气候变化导致藻类生理状态变化的生物方法(Ahn et al., 2019;Ismail等人,2022),并用于确定营养状况(Kim等人,2020)。在全球范围内,最近的研究集中在水体特征的变化与浮游植物生化组成的变化之间的关系。然而,到目前为止,在埃及只有少数研究报告了这些主题。研究站浮游植物的生化组成呈现出显著的空间差异(图5),这可能与浮游植物种类和理化参数的差异有关(Kim et al., 2022)。此外,藻类生长阶段在决定其大分子含量变化方面起着重要作用(Fernández-Reiriz et al., 1989)。从EH、SG、G和AQ站收集的浮游植物中,蛋白质是主要的生化成分,其次是碳水化合物和脂类,它们通常反映了充足的氮条件和生理健康的浮游植物,相对于生产区域具有较高的相对生长速度(Kim et al., 2022)。在其他站点,估计的CHO (av. 232.23µg L−1)是M、B、RB和PS站点浮游植物生物量中最丰富的成分,PRO (av. 215.74µg L−1)是第二贡献,这表明这些站点浮游植物生长的氮饥饿(Danovaro等,2000)。此外,Fathi等人(2005)指出,CHO的增加是由污染地区藻类PRO的下降引起的。
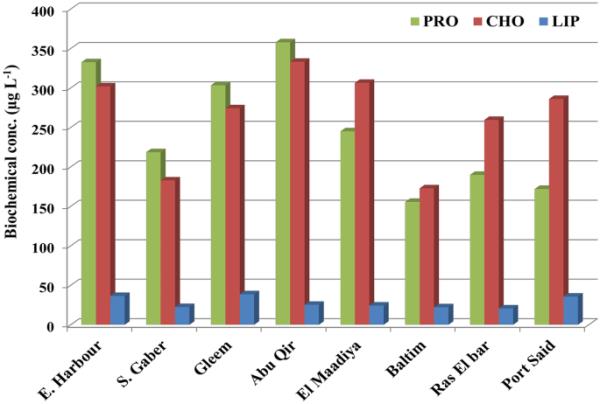
东地中海亚历山德里亚沿岸不同海岸浮游植物的生化含量(µg L−1)
浮游植物总脂含量(28.13µg L−1)较小(图5)。最大LIP分别在g、EH和PS处检测到38.6、36.4和35.6µg L−1。检测到的大分子含量高于Ismail等人(2022)在EH报告的2020年期间的结果。
蛋白质达到三个主要峰;AQ含量最高(357.77µg L−1),种类最多的是生毛角毛藻(Chaetoceros)和肉芽草(Melosira granulata);硅藻占高峰日(63 × 103细胞L−1,Chl为0.52µg L−1)。第二个峰(303.33µg L−1)出现在g站,伴随着透明质杆菌丰度的开始(52 × 103细胞L−1,Chl为0.65µg L−1)。由于蛋白质是含氮化合物,浮游植物群落蛋白质含量与水中总氮含量呈显著相关(r = 0.697) (Yun et al., 2015)。蛋白质的增加被认为是浮游植物代谢中不存在氮胁迫的指标(Ahn et al., 2019)。
收集到的浮游植物的蛋白质和碳水化合物比例从西向东递减,这表明浮游植物的生长受到氮限制(Danovaro et al., 2000)。该比率已被用于表征n条件和细胞的生理状态(Ismail等人,2022;Kim et al., 2015)。在N-饥饿条件下,CHO和LIP都是积累的,因为它们不需要N,作为储存材料和结构成分,它们可能是一种生存的反应机制(Hu, 2004)。
虽然关于水参数对浮游植物群落大分子含量的直接影响的数据有限,但已知它们是藻类生长所必需的。由于减少或/和增加,可能会减少或/和刺激生化含量的产生(Boëchat & Giani)。, 2000)。通过相关分析,本研究中对浮游植物“CHO和LIP”生化含量影响最显著的因素是温度(表2S),而水温是影响微生物群落生理活性及其生长的重要非生物因素(Zhao et al., 2020)。温度升高会抑制PRO合成(r = - 0.67),从而导致藻类生长速度降低(Schulte, 2015)。此外,温度对LIP (r = - 0.596)和CHO含量(r = - 0.641)也有负面影响,如Zhao et al.(2020)先前所述。盐度对水生生态系统中浮游植物的生化含量(PRO (r = 0.441)和CHO (r =−0.421))也有影响。Kumar和Saramma(2018)也发现了类似的现象,他们发现随着盐度的增加,PRO含量增加,CHO含量降低。此外,PO4对CHO (r = 0.558)和PRO (r = 0.467)有正向影响,而磷是藻类细胞CHO和PRO代谢途径中必需的营养物质(Yaakob, 2021)。PRO、总氮(r = 0.691)和NO3 (r = 0.787)的变化趋势相同,而氮是浮游植物生长的第二大营养物质,缺氮期间PRO的相对下降(Yaakob et al., 2021;Kim et al., 2022)。另一个影响大分子含量的因素是DO浓度。Boëchat和Giani(2000)证明,在低DO水平下,浮游植物细胞似乎降低了其蛋白质含量(r = 0.708)。另一个可能对生物化学成分产生重大影响的因素是浮游植物的组成。不仅是藻类种类,还有生长阶段(Boëchat & Giani, 2000)。硅藻与CHO (r = 0.425)和LIP (r = 0.406)均有关系,而硅藻细胞的主要成分是脂质,硅藻中平均脂质含量可达干重的25% (Levitan et al., 2014)。此外,它们还会产生大量的细胞外聚合化合物,如糖蛋白和多糖(Haynes et al., 2007)。
下载原文档:https://link.springer.com/content/pdf/10.1007/s10661-023-11690-z.pdf




